支付宝花呗额度换现技巧,方法总结花呗如何自己刷出来
|
○ 昆虫的嗅觉受体是一种离子通道。当气味分子与嗅觉受体结合时,离子通道打开,离子内流,引起动作电位。○ 研究人员研究了石蛃的嗅觉受体,发现不同的气味分子与受体的同一口袋对接。这不是经典的锁钥模式,而是一种一对多的结合方式。 ○ 他们发现即使是改变受体口袋中的单个氨基酸,也足以改变受体口袋的结合特性。这可能解释了为什么昆虫的嗅觉受体演化得如此之快,且物种间的差异如此之大。 撰文| Jordana Cepelewicz 编译|阿朔 审校|Lixia 编辑 | Jiahui,EY 嗅觉(而非视觉),是大多数动物最重要的感官。它使动物们可以找到食物,躲避危险以及吸引配偶;它统摄动物的感知并引导其行为;它决定了动物如何解释和应对周围海量的感官信息。 然而,嗅觉也可能是我们的感官中最难以理解的,部分原因在于它的输入信息的复杂性。我们认为的单一气味——清晨的咖啡味、夏日暴风雨后的湿草味、洗发水味或香水味——通常是数百种化学物质的混合物。对于动物来说,要探测和辨别对其生存至关重要的多种气味,其嗅感觉神经元上有限的受体必须以某种方式识别大量的化合物。因此,单个受体必须能够对许多不同的、看似无关的气味分子作出反应。现在,在阐明嗅觉过程的起始阶段方面,新的研究已经迈出了至关重要和备受期待的一步。在今年早些时候在线发表的预印本中,洛克菲勒大学的研究小组首次提供了嗅觉受体与气味分子结合时的分子视图。瑞士洛桑大学的生物学家理查德·本顿(Richard Benton,没有参与这项新研究)说,自从30年前发现嗅觉受体以来,“这一直是该领域的一个梦想”。 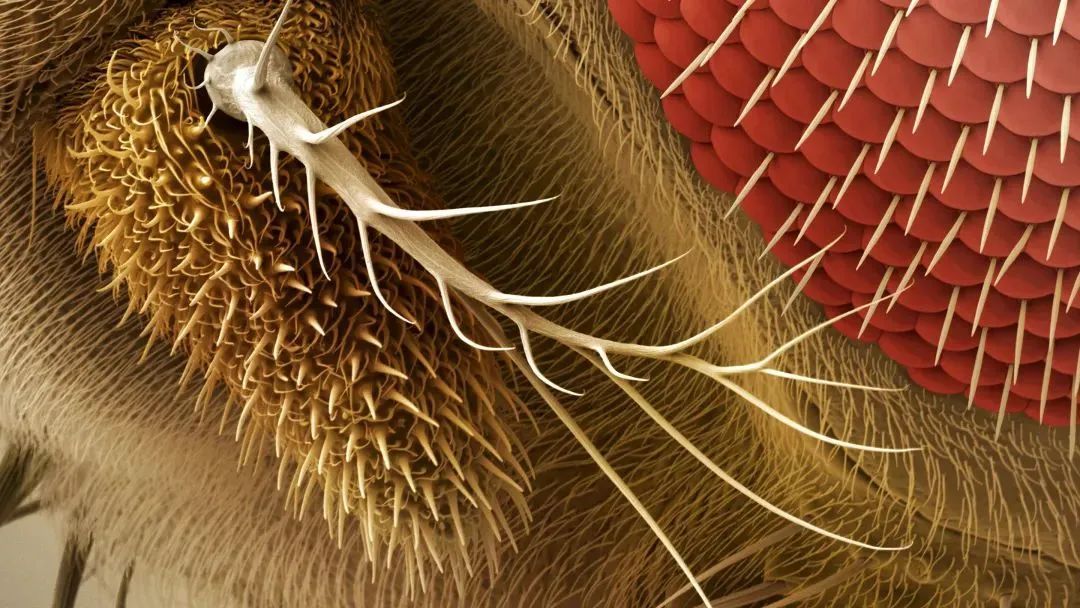 果蝇的触角。丨图片来源:Science Image of Fruit fly antenna from PS MicroGraphs 这个结果对确认动物如何识别和区分海量的气味有很大帮助。它还阐明了受体活性的关键原理,这可能对了解化学感知的演化、理解其他神经系统和过程如何工作,以及开发诸如靶向药物和驱蚊剂等具有深远意义。有几个假说争相解释嗅觉受体如何达到必要的柔性。一些科学家提出,受体对气味分子的单一特征做出反应,比如形状或大小;然后大脑可能通过综合来自不同受体的信息识别出一种气味。其他研究人员认为,每个受体都有多个结合位点,可与不同种类的化合物对接。但为了弄清楚这些观点中哪个是正确的,他们需要看到受体的实际结构。 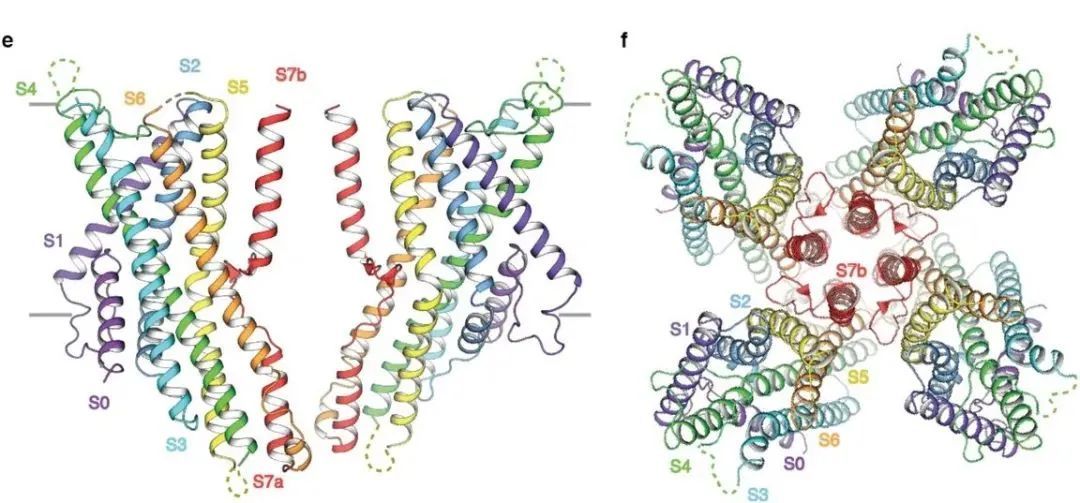 从侧面(e)和顶部(f)显示的 MhOR5 (石蛃嗅觉受体)的冷冻电镜结构。丨图片来源:https://doi.org/10.1101/2021.01.24.427933 原始昆虫的嗅觉受体 洛克菲勒研究小组将研究方向转向了石蛃的受体相互作用。石蛃是现存最原始的昆虫,有一个特别简单的嗅觉受体系统。在昆虫中,嗅觉受体是一种离子通道,当气味分子与它们结合时,它们就会被激活,从而引起动作电位。在世界各地的昆虫物种中有数百万种这样的离子通道,因而嗅觉受体可能是自然界中规模最大、最多样化的离子通道家族。因此,它们必须小心地平衡共性和特异性,既要有足够的柔性来检测大量的潜在气味,又要有足够的选择性来可靠地识别重要的气味(这些气味可能因物种或环境的不同而存在显著差异)。  研究人员在石蛃(jumping bristletail)身上发现了一种气味受体。这种无翅昆虫的嗅觉系统更简单、更原始,因此是理想的测试对象。丨图片来源:Yasunori Koide 是什么机制让它们能够把握如此恰好的方向,并以这种方式演化?用传统的确定蛋白质三维分子结构的方法来研究嗅觉受体,效果并不理想。在这些方法所需要的条件下,嗅觉受体往往会折叠错误、行为异常或变得难以分辨。但最近的技术进步,尤其是一种称为冷冻电子显微术(cryo-electron microscopy)的成像技术,使研究者们得以尝试新的方法。他们研究了三种不同构形下石蛃嗅觉受体的结构:一种是受体本身,一种是与常见的气味分子丁香酚(eugenol,闻起来像丁香)结合,一种是与驱虫剂避蚊胺(DEET)结合。然后,他们将这些结构进行了比较,细微至单个原子,以了解气味结合是如何打开离子通道的,以及单个受体是如何确认形状与大小都有显著差异的化学物质的。 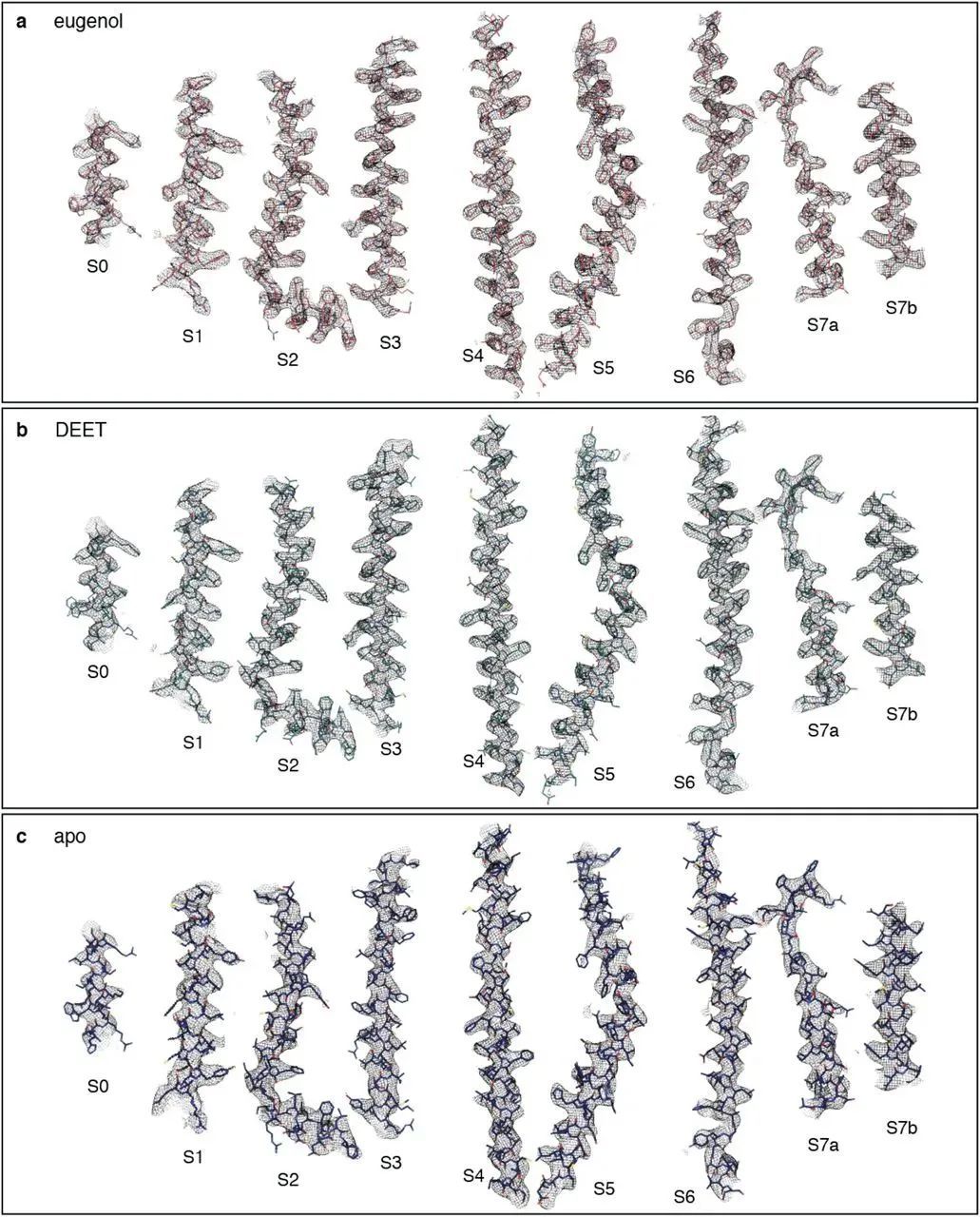 建模区域的冷冻电镜密度,从上至下:丁香酚结合结构,避蚊胺结合结构,蛋白质的结合前结构。 研究人员发现,尽管作为分子,避蚊胺和丁香酚没有太多共同之处,但它们都与受体的同一位置对接。该位置是一个深而简易的口袋,内侧排列着许多氨基酸,有利于形成松散的弱相互作用。丁香酚和避蚊胺利用不同的相互作用在口袋内驻留。进一步的计算模拟显示,每个分子能以许多不同的方向结合,而且许多其他种类的气味化合物也可以以类似的方式与受体结合。这不是一对一的锁钥模式,而是一种一对多(one-size-fits-many)的方法。(见段末示意图)嗅觉受体“正在对分子进行更全面的识别,而不仅仅是检测任何特定的结构特征。”这项研究的作者瓦妮莎·鲁塔(Vanessa Ruta)说,“这是一个非常与众不同的化学逻辑。”当鲁塔和她的团队改变受体口袋时,他们发现即使是单个氨基酸的突变也足以改变其结合特性。而这又足以影响受体与许多化合物的相互作用,完全重置受体所起作用的对象。例如,扩大受体口袋,增加了它对避蚊胺(较大分子)的亲和力,而降低了它对丁香酚(较小分子)的亲和力。这可能是因为丁香酚尺寸较小,无法很好契合较大的受体口袋)。这样的变化也会对受体更广泛的气味检测“调色板”(odor-detecting palette)产生下游效应,而研究人员并没有确定这些影响。该小组的观察可能解释了为什么昆虫的嗅觉受体演化得如此之快,在物种间的差异如此之大。每一种昆虫可能都演化出了“自己的独特受体,这些受体非常适合其特定的化学生态位。”“这告诉我们,除了受体与一堆配体弱相互作用之外,还有更多的事情在发生。”神经生物学家鲍勃·达塔(Bob Datta)说。一种建立在单一结合口袋周围的受体,其反应剖面(response profile)可以通过微调来调整。如果更广泛地改变该受体的化学成分,可能会加快演化的进程。该受体的结构也证实了这一观点。鲁塔和她的同事发现,受体由四个蛋白质亚基组成,松散地结合在通道的中心孔上,就像一朵花的花瓣。随着受体的多样化和演化,只有中心区域是保守的;而控制其余受体单位的基因序列较少受到限制。这种结构组织意味着受体可以适应广泛的多样化。这种受体水平上的轻微演化限制可能会对下游的嗅觉神经回路施加巨大的选择压力:神经系统需要良好的机制来解码受体活动的混乱模式。“实际上,嗅觉系统已经演化到采取任意的受体激活模式,并通过学习和经验赋予它们意义。”鲁塔说。 |

 支付宝扫一扫
支付宝扫一扫 微信扫一扫
微信扫一扫












评论列表